
A dividing cancer cell
A Cancer cell is a pale cast of thought, pregnant with desire, packed with motive and reasoning more murderous then all ill-intent!
It was a cool dark night. Not a chirp from a bird or a howl from an owl. The trees were still. The flowers limp with dew. Not a soul inhabited the streets, save one dark shadow. The face masked and with gloves to cover all skin only the windows to his cloaked soul shone through. He weaved in and around the trees after stepping out of his bungalow; lurking behind the walls of other homes, seeking opportunity and stealth. He let out a soft bird-call that was answered immediately from beyond and another shadowy figure emerged and as quickly disappeared into the thicket of darkness. The game was afoot. The object of their desire was the wealth in one lone house perched amidst the weeping willows and the might oaks. That house was silent. The dog lay sleeping in its little abode guarding only its midnight dreams. One of the robber crossed the street and with speed and an artful craft making nary a sound, he jumped the fence with ease, crossed over and threw a half-eaten steak in front of the dog that barely stirred, to rob him off his scent. He then proceeded carefully, using the dexterity of his fingers and the learned manipulation of a simple right-angled tool and opened the multi-pronged door lock. A hiss sounded and then a low tone. The place was alarmed. The other robber appeared from beyond the shadows and moved to the opposing wall panel that housed the brains of the alarm system and using a cell phone and two wires hijacked the system into quiet obedience. Again with light-footed efficiency and agility one walked straight to the library, unhinged the gold-framed photo of the owner and behind it exposed the safe – the heart of the owner’s life-long enterprise. The safe was no match to his ingenuity. It opened without a squeal or clamor. The other robber surreptitiously and without so much as a sound gathered other valuables. A life was undone.
Imagine the cloaked robbers as cancer cells. It is the same sociopathic behavior of ingratiation without lack of empathy for the host and a constant self-perpetuating desire. That same motivation feeds both. It is how cancer spread (metastasizes) occurs. Surprised? You shouldn’t be. Let us take a short journey into the fledgling field of metastasis.

The cancer cell, like the masked, cloaked robber uses similar stealth and tact to circumvent the host defense, for there are many hurdles that favor the normal organs against the spread of disease and all have to be neutralized one by one. [Italics represent quotes from various studies listed below in references]



Spread: A. Primary tumor, B. Local invasion E, Lymphatics D. Blood vessel, F. Fascia
First Hurdle: The cancer cell has to grow and multiply. For this, it co-opts the vascular system (blood supply) and releases hormones in its environs. It also, through communications within the normal surrounding community of cells via cross-talk, establishes a quid-pro-quo relationships in the form of, “I am here, you are here, lets be friends” kind of way.

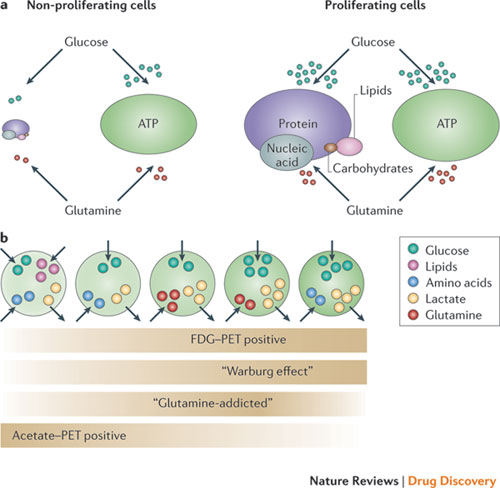
Soon having established the groundwork for attaining the nutrients for itself to grow, it manipulates its neighbors into transitioning from their normal functioning selves into cells that unknowingly and inadvertently help in the choreographed dance of stealth and duplicity. There is a close relationship between the metabolic needs of the cancer cells and that of the host micro environment. Not only do the metabolic needs of the cancer cell but those of the host human plays a significant role in perpetuation and progression of this disastrous of all maladies, they also enhance or suppress it:

Soon having established the groundwork for attaining the nutrients for itself to grow, it manipulates its neighbors into transitioning from their normal functioning selves into cells that unknowingly and inadvertently help in the choreographed dance of stealth and duplicity. There is a close relationship between the metabolic needs of the cancer cells and that of the host micro environment. Not only do the metabolic needs of the cancer cell but those of the host human plays a significant role in perpetuation and progression of this disastrous of all maladies, they also enhance or suppress it:

“Genetic alterations that are associated with cancer often occur in these same signalling pathways, which suggests that both environmental and genetic factors influence the metabolic heterogeneity that is present across tumours… Indeed clinical studies have linked altered whole-body metabolism to cancer development, progression and poor treatment outcomes. Indeed, obesity, hyperglycaemia and insulin resistance are all associated with an increased risk of developing cancer and are associated with worse clinical outcomes in patients with cancer… Increased circulating levels of insulin and insulin-like growth factor (IGF) have been linked with cancer pro-gression, which suggests that obesity and insulin resistance promote cancer at least in part by activating signalling pathways that drive cell growth. These same signalling pathways also drive nutrient uptake into cells and regulate enzymes in glycolysis, which implies that hormonal changes can have important indirect effects of cancer. Furthermore, elevated levels of glucose alone may promote increased glucose uptake in some cells, and lower circulating levels of glucose are associated with better cancer treatment outcomes”
The well known Warburg effect after Otto Heinrich Warburg, who hypothesized that the cancer cell needs two pathways for survival: the glycolytic pathway whereby the cell breaks down the glucose to obtain energy and the Lactic Acid pathway, whereby the cancer cell needs to eliminate the excess byproduct of glucose consumption (waste product), is alive and well. Targeting both sides and not affecting the normal cell is a challenge that consumes many in the field of oncology.
In the same context, Insulin Growth Factors (IGFs) are shown to increase tumor proliferation by activating the phosphoinositide 3-kinase (P13-K) pathway. This pathway is affected by the nutrient availability and the pathway signaling inhibition by drugs downstream can be impeded by the excess of nutrients, thus the signal once impeded by such an overabundance of resource, may not reach the target inside the nucleus to initiate growth. An agent that seems to influence the m-TOR, a downstream path, of the P13-K pathway is a drug called, Rapamycin. However in the presence of excess nutrients, such as, glucose the efficacy of Rapamycin is compromised. It is thus important for the human host not to indulge in food and therefore obesity. Excess food and thus obesity are confounding factors that help the robbers rob!

In a recently discovered entity dubbed the Epithelial-Mesenchymal-Transition or EMT, the E to M phase transition can also go from M to E (MET) and indicates the fluid flow of cells morphing by desire and motivation, by acquiring properties of others that have better abilities to migrate, invade and spread. All of these phase transitions within the cancer environment occur for the sole purpose of mitigating any difficulties for escaping the firewalls erected by the basement membranes and other fascia as defenses within the host.

Some cancers like the alveolar cell carcinoma of the lung and the clear cell carcinoma of the ovary have a field day in “spreading” due to their proximity to an “open-field.” Both types of cancers also manipulate various cytokines and enzymes and by promoting such, they are able to modify the epigenetic system to abrogate the tumor-suppression algorithms within the DNA code from coming on board. The Phase transition, EMT is an aberrant activation of a latent embryonic Program encoded within the cells. This very deterministic behavior lends to self-renewal capability, antigenic expression and later, resistance to therapy and is the object of recent scrutiny.

Some cancers like the alveolar cell carcinoma of the lung and the clear cell carcinoma of the ovary have a field day in “spreading” due to their proximity to an “open-field.” Both types of cancers also manipulate various cytokines and enzymes and by promoting such, they are able to modify the epigenetic system to abrogate the tumor-suppression algorithms within the DNA code from coming on board. The Phase transition, EMT is an aberrant activation of a latent embryonic Program encoded within the cells. This very deterministic behavior lends to self-renewal capability, antigenic expression and later, resistance to therapy and is the object of recent scrutiny.

“It is increasingly acknowledged that aberrant activation of a latent embryonic program - known as the epithelial-mesenchymal transition (EMT) - can endow cancer cells with the migratory and invasive capabilities associated with metastatic competence [1-3]. Moreover, several lines of evidence have converged in recent years to support the notion that not all cancer cells within a given tumor are equal in terms of their tumor-initiating potential. The emerging paradigm posits that tumor progression is driven by a small subpopulation of cancer cells - termed cancer stem cells (CSCs) or tumor-initiating cells - that exhibit two defining characteristics: the ability to self-renew and the ability to regenerate the phenotypic heterogeneity of the parental tumor [4]. CSCs have thus been implicated both in initiating and sustaining primary tumor growth and in driving the seeding and establishment of metastases at distal sites… During normal embryonic development, EMT serves to loosen cell-cell contacts and to enhance intrinsic cell motility, thus paving the way for the extensive cell movements required for gastrulation and organogenesis… EMT can be induced by a plethora of extracellular stimuli, including hepatocyte growth factor, epidermal growth factor, platelet-derived growth factor, Wnt, Notch, Sonic hedgehog and transforming growth factor beta (TGFβ) as well as components of the extracellular matrix such as collagen and hyaluronic acid and adverse conditions such as hypoxia [1,3]. These diverse stimuli trigger a multitude of signal transduction pathways that converge on several EMT-inducing TFs, including Snail, Slug, Zeb1, Zeb2, Twist, FoxC2 and Goosecoid, many of which are frequently overexpressed in breast cancers”
The breast, it may be reminded, is an organ worthy of considerable study because it undergoes expansion and then involution for each pregnancy cycle and thereby provides us with an indirect indication of the presence of breast stem cells that undergo morphogenetic branching via many transcriptional factors during these single or multiple pregnant phases. Additionally, a family of Metastatic Tumor Antigens (MTA1-3) proteins that are also involved in the spread of breast cancer:
“The metastasis tumor antigen (MTA) family of proteins, MTA1, MTA2, and MTA3, are components of chromatin remodeling pathways with potential roles in breast cancer.”
Using the genetic profiles of breast cancers, six different subtype populations of differing molecular signatures have been identified; luminal A, luminal B, HER2+, basal-like and claude-like. Most chemotherapy drugs attack the cancer cells but do not address the de-novo or phase transitioned cancer stem cells that have evolved. An anti diabetic drug “Metformin” seems to address that directly by killing Cancer Stem Cells (CSCs). Now, that might be a direction towards cure should further studies confirm this violate action as being a true representative of going after the “culprit!” Imagine destroying the menagerie of propagating cancer cells with chemotherapy, targeted biologic therapy and then going after the sociopath itself! Thus based on the predicate, which stipulates that the primary tumor is governed by different sets of molecular rules from those of the metastatic cancer elucidates the realm of differential therapy with multiple “medical” and “surgical” scalpels. Better yet, if one can choke off the system completely without affecting the normal cells remains the holy-grail! Refining the sociopath and changing his or her motivation is a daunting path since it is difficult to determine who is who. It is a difficult task indeed. But understanding the needs and depriving the aggressor may be the simplest preventative measure towards success. We might just have to live by “Dunkin Broccoli rather than Donuts.”

Got you out of the comfort zone, didn’t I? Sorry. Let us walk a little slower. Smell the fresh air and take a deep breath once again. The entire orchestral symphony played by the cancer cell is a strategy for self-renewal and thus self-perpetuation. The cancer cell wants to survive and propagate. That is the code writ in its code. It thus manipulates its environs to suit its own needs. Just like the unmasked, and uncloaked man is a perceived normal functioning member of his household and of the larger community, yet lurking within him is the unseen evil betrayal. No one is aware, or even concerned that this mild-mannered sociopath is a burglar, aiding and abetting others to join his league. Acquiring these traits of using enzymes to cut through the jungle of basement membranes, fatty tissue, fascia and a dense arborized collection of capillaries and supporting cellular network, is like the prince hacking at the forest in search for Snow White, only this is not the prince but the witch using her unlimited powers to dissolve and destroy so she can govern and control the purity. The cancer cell makes its escape by recruiting host micro environment features, such as the Tumor Associated Macrophages or (TAMs) to help make the break through the jungle. TAMs have the ability to secrete various dissolving enzymes to clear the path for the progress of the cancer cell and confuse its own immunity police. Macrophages thus become the unwitting accomplices in the robbery.

Primary cancer, CTCs and distant Metastasis

Main Lymphatic System
The Main lymphatic system originating in the abdomen also ends up in the vascular system eventually, it is called the Cisterna Chylii and dumps its lymph via the thoracic duct into the large subclavian venous system on the left side of the neck. This is why there is a revitalized interest in the CTCs (Circulating Tumor Cells). The presence of the CTCs is object of renewed interest to try to detect their presence and decipher their vulnerabilities. 40% of all comers with breast cancer have spread of cancer cells in the bone marrow and an equal number of lung cancer in early stage I state have cancer cells in the pleural sac that lines the lung. No wonder there is recurrence in early stage disease at a later date. The “cat had already climbed out of the bag.”

One can detect their (CTCs) (cancer cells) genomic make up and fire targeted therapies at them in an attempt to cure. Although it may seem that the cancer cell has it all in its favor. Not so! Another hurdle for the free-floating CTCs in the blood-stream is that having lost cell to cell adhesion, in their free-floating mode, they rapidly undergo anoikis (apoptosis or cell death) When observed morphologically, it is unknown if these CTCs are alive, playing possum or just dead and effete waiting for the dirt heap. The cancer cell “Mission Impossible” smart, adaptive and self-protective, capability has devised elaborate special anti-apoptotic mechanisms by expressing Bcl-2, Bcl-XL or FAK genes, to keep itself alive from death till it finds a host tissue organ. The self-destruct message is not invoked that easily. No poof! Only more smoke and mirrors. Some of these cells may live in the body undetected for months to years and resurface again. It is well known that years later the same cancer, considered cured, can raise its ugly head and new molecular data seems to shed some light on this phenomenon. The argument remains whether it is the cell that clicks its machinery into action or the abrogation of responsibility by a taxed immune system that allows such spread, no one is certain just yet.
Third Hurdle: From the blood vessels migrating back into an organ is another deterrent for the cancer cell. There are two issues with this hurdle: One, for the cancer cell to re-invade the endothelial and muscular layers of the blood vessel wall to reach the outside reference of the vascular system is difficult and, two, to find a location where the cell is comfortable in its existence. (Someone who loves the beach will want a beachfront property and those desiring mountaintop abodes will find their destinies). So there is a “soil” issue that remains to be resolved. Oh yes these cancer cells are picky! The cancer cell has protrusions on its surface that look for target sites on the organs. For instance colon spreads to the lymph nodes and the liver, breast cancer spreads to the lymph nodes and the bones, prostate cancer spreads mainly to the bones. All these predilections are based on the “homing devices” the cancer cells carry on their surfaces. Not to say that the cancers do not deviate and attack other organs, they do, but the majority, behave in the fashion elicited for a reason. A transcription factor 3-gene being an adaptive-response gene may serve to integrate stromal signals from the tumor micro environment and in so doing bring the invader to its selected host site. Or in other words the signals from the host organ are received and processed by the cancer cell and appropriate obfuscating response is given to the host to confuse and thus allow easy access. (It is akin to throwing a steak to the dog). In Osteosarcoma (A sarcoma (cancer) of the bone) where 20% of patients have metastases at the time of initial diagnosis and 40% in later stage, 80% of the metastases are to the lung! Thus a special proclivity to spread to the lung tissue exists in osteosarcoma cancer cell.
“The metastatic behaviour of OS (Osteosarcoma) is very distinct as over 80% of all metastases arise in the lungs and other organs usually remain unaffected. This suggests that the circulating tumour cell specifically ‘homes’ to distinct molecules that are expressed on the endothelium of the organ of preference”
Fourth Hurdle: Once inside the organ, the cancer cell has to fight off the natural defense of cellular density (cramped quarters) within the organ of its desire. Again the EMT and MET come into play at the host site.
“A critical molecular event underpinning the dissolution of cell-cell contacts during EMT is the loss of E-cadherin, a key component of adherens junctions… The loss of E-cadherin releases β-catenin into the cytosol and elicits activation of the canonical Wnt signaling pathway [1,3]. Moreover, the impairment of E-cadherin function, together with the downregulation of components of tight junctions and desmosomes (for example, claudins, occludins, desmogleins and desmocollins) and polarity genes, contributes to the dissolution of inter-cellular contacts and the loss of apico-basal polarity… The ensuing reorganization of the actin cytoskeleton and the intermediate filament network and the acquisition of front-back polarity dramatically alter the cellular architecture, while the secretion of extracellular matrix components and matrix metalloproteinases remodels the extracellular matrix. Collectively, these changes disrupt the contiguity of the tissue epithelium and render the cells intrinsically able to migrate - independent of one another - and to invade the underlying stromal compartment by breaching the basement membrane”
It does so by “talking” to its neighbors and coercing them into allowing residence and ultimately though vascular fiat of co-opting the vascular endothelial factor release it finds itself a stream of palatable nutrition (Employing the same previously well-managed method). Cancer cells also block the immune network by sending alternate and confusing signals to the immune cells, (T and B lymphocytes, Dendritic Cells and the macrophages). While the first robber is decoding codes of protection, the second robber is sighting, gathering up the valuables and laying bare the foundations of an existence.

Dendritic Cell
“Tumour cells, either circulating or at the site of metastases, can modulate the immune system of the host in order to achieve a survival advantage. Down-regulation of cell surface receptor HLA class 1 is one of such mechanisms. This impairs the recognition of tumour cells by the host cytotoxic T-lymphocytes. Tumour cells can also induce the production of immunosuppressive molecules such as IL-10… Fas also plays a role in immune evasion. Fas expression leads to recognition by, and activation of cytotoxic natural killer (NK) cells and promotes elimination from the circulation by the host immune system.”
Additionally they send out alarms of inflammatory signals throughout the blood system. (IL18, NFkB, TNFa etc.) These cytokines have the ability to modulate the genetic makeup of the immune cells by bringing them in full force to bear on the enemy (in this case the cancer cell) and then by disinformation and direct assault causing attrition forcing the defensive host army to laying down the arms and capitulating. Thus the cancer cell kills the immune-surveillance for the single and simple act of self-perpetuation.

Hypoxic (low oxygen) environments are not conducive to most cells. Metastatic cancer cells reap the rewards in a low oxygen environment where they find themselves frequently, both in the initial phases when the framework of blood vessels have not yet been established and late stages of growth, where there are too many cancer cells in the center of the tumor where the co-opted blood vessels cannot reach by distilling out HIFs. They use hypoxia as a mediator to release (Hypoxia Inducible Factor) s HIFs to force other signaling proteins to modify the DNA to build more bridges over breached dams for their own survival.
“Low oxygen tension in the primary tumour is associated with metastasis in soft tissue sarcoma, cervix carcinoma and carcinoma of the head and neck. Multiple mechanisms may be involved in hypoxia-induced metastasis. Thus, hypoxia followed by reoxygenation may induce point mutations and DNA strand breakage leading to deletions, amplifications and genomic instability…Moreover, hypoxia may induce a temporary increase in the expression of gene products involved in the metastatic cascade, either through gene amplifications or through normal physiological processes by activating oxygen sensors, hypoxia signal transduction pathways and DNA transcription factors”
Every line of defense proposed by the host is subject to mitigation and denial of sustainability by the cancer cell. This co-opting mechanism is the hard and tedious work of Darwin’s old adage, “The survival of the fittest.” The cancer cell having acquired devious means, wants to survive. The cancer cell is unfortunately better equipped since it has understood the molecular machinery of survival better then the host has determined new adaptive lines of defense. There is more tit than tat. The fight between the host and its cargo of renegades, is a battle of intense planning, scrutiny, subterfuge and all out assault that rages in deft silence.
Therapeutic Options for metastases exist currently in the form of surgery (metastatectomy), radiation therapy, chemotherapy and biologic therapy. The former is surgical removal of the metastatic sites. There is proof at least in Osteosarcoma, and in Colon cancer where known only-metastatic cancer sites when removed (osteosarcoma metastases to lung and colon cancer metastases to the liver (in colon cancer, as long as no intervening spread or in-transit spread into lymph nodes has occurred)) yield better survival. It is also well known that patients with concurrent metastases have a worse survival then sequential ones. The precept being that the cancer cell has already achieved multiple DNA transgressions in the former and acquires them later in the latter. The steps needed for metastases to occur in osteosarcoma specifically are detailed in the table below: (listed in references):
Steps of metastasis | Molecular involvement |
Migration and invasion | MMPs |
m-Calpain | |
Wnt | |
Src | |
Notch | |
(a) Anoikis resistance | PI3K/Akt |
Src/PI3k/Akt | |
Src/Ras/MAPK | |
NF-κB | |
Wnt/β-catenin | |
BcL family | |
(b) Apoptosis resistance | Src |
NF-κB | |
Wnt/β-catenin | |
Fas/FasL | |
Evasion of immune system | HLA-1 |
IL-10 | |
Fas | |
Arrest and extravasation | CXCR4-CXCL12 |
CXCR3-CXCL9-11 | |
CXCR4/MMPs | |
CXCR3-4/Erk/NF-κB | |
Adherence | Ezrin/MAPK/Akt |
Ezrin/β4-Integrin/PI3K | |
CD44/Akt/mTOR | |
Dormancy | Integrin-α5β1 |
Integrin-α5β1/Erk/p38 | |
Bcl-XL | |
IGF/PI3K | |
ECM | |
Angiogenesis and proliferation | EGFR. PDGFR, VEGF, IGFR, TGF-β |
MMPs | |
VEGF/Erk/NF-κB | |
VEGF/PI3K | |
EGFR/Src/Ras/MAPK/STAT3 | |
Src | |
Integrin/PI3K/Erk1-2 | |
Wnt/β-catenin/CyclinD-Survivin |
Understanding of this elaborate and intricate fabric woven together by a rag-tag group of wayward cells is the difference between life and death. The matters remain intricate and more and more complexity continues to be added to this heavy tome of discussion. The answer may lie at fundamental levels. There may be a final common pathway, or as we now know, there may be many layers to the finite bottom of this “rabbit hole.” Each step and each experiment brings us closer and sometimes inadvertently leads us further from the truth through the minutia of “discussions.” We will uncover that truth eventually and relegate it at some intermediate level of a “chronic disease’ and eventually to the vast heap of “past maladies.” Whenever the end-result, the journey is far from over and ripe for the taking.
References:
Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009;139:871–890
Prat A, Parker JS, Karginova O, Fan C, Livasy C, Herschkowitz JI, He X, Perou CM. Phenotypic and molecular characterization of the claudin-low intrinsic subtype of breast cancer. Breast Cancer Res. 2010;12:R68
Polyak K, Weinberg RA. Transitions between epithelial and mesenchymal states: acquisition of malignant and stem cell traits. Nat Rev Cancer. 2009;9:265–273
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, Campbell LL, Polyak K, Brisken C, Yang J, Weinberg RA. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008;133:704–715
Dontu G, Al-Hajj M, Abdallah WM, Clarke MF, Wicha MS. Stem cells in normal breast development and breast cancer. Cell Prolif. 2003;36(Suppl 1):59–72
Fata JE, Werb Z, Bissell MJ. Regulation of mammary gland branching morphogenesis by the extracellular matrix and its remodeling enzymes. Breast Cancer Res. 2004;6:1–11
Hirsch HA, Iliopoulos D, Tsichlis PN, Struhl K. Metformin selectively targets cancer stem cells, and acts together with chemotherapy to block tumor growth and prolong remission. Cancer Res. 2009;69:7507–7511
Creighton CJ, Chang JC, Rosen JM. Epithelial-mesenchymal transition (EMT) in tumor-initiating cells and its clinical implications in breast cancer. J Mammary Gland Biol Neoplasia. 2010 Jun;15(2):253-60. Epub 2010 Mar 31.
Geiger TR, Peeper,D. Metastasis mechanisms. S Biochim Biophys Acta. 2009 Dec;1796(2):293-308. Epub 2009 Aug 14.
.
Barbour A, Gotley DC. Current concepts of tumour metastasis. Ann Acad Med Singapore. 2003 Mar;32(2):176-84.
Vander Heiden, M. G., Cantley, L. C. & Thompson, C. B. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 324, 1029–1033 (2009)
Harting MT, Blakely ML. Management of osteosarcoma pulmonary metastases. Semin Pediatr Surg. 2006;15(1):25–29.
Krishnan K, Khanna C, Helman LJ. The biology of metastases in pediatric sarcomas. Cancer J. 2005;11(4):306–313.
Lafleur EA, Koshkina NV, Stewart J, et al. Increased Fas expression reduces the metastatic potential of human osteosarcoma cells. Clin Cancer Res. 2004;10(23):8114–8119.
Rubin EM, Guo Y, Tu K, et al. Wnt inhibitory factor 1 decreases tumorigenesis and metastasis in osteosarcoma. Mol Cancer Ther. 2010;9(3):731–741.
J. PosthumaDeBoer, M. A. Witlox, G. J. L. Kaspers, and B. J. van Royen. Molecular alterations as target for therapy in metastatic osteosarcoma: a review of literature. Clin Exp Metastasis. 2011 June; 28(5): 493–503.
Caitlin D May, Nathalie Sphyris, Kurt W Evans, Steven J Werden, Wenjun Guo and Sendurai A Mani1 Epithelial-mesenchymal transition and cancer stem cells: a dangerously dynamic duo in breast cancer progression. Breast Cancer Res. 2011; 13(1): 202.
Zhang H, Stephens LC, Kumar R. Metastasis tumor antigen family proteins during breast cancer progression and metastasis in a reliable mouse model for human breast cancer. Clin Cancer Res. 2006 Mar 1;12(5):1479-86.
Rofstad EK. Microenvironment-induced cancer metastasis. Int Radiat Biol, 2000 May;76(5):589-605.
Qian, B., Y. Deng, J. H. Im, R. J. Muschel, Y. Zou, J. Li, R. A. Lang, J. W. Pollard. 2009. A distinct macrophage population mediates metastatic breast cancer cell extravasation, establishment and growth. PLoS One 4: e6562
Bingle, L., N. J. Brown, C. E. Lewis. 2002. The role of tumour-associated macrophages in tumour progression: implications for new anticancer therapies. J. Pathol. 196: 254–265.
Joyce, J. A., J. W. Pollard. 2009. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 9: 239–252.
Lewis, C. E., J. W. Pollard. 2006. Distinct role of macrophages in different tumor microenvironments. Cancer Res. 66: 605–612.
Matthew G. Vander Heiden Targeting cancer metabolism: a therapeutic window opens Nature Reviews Drug Discovery 10, 671-684 (September 2011) | doi:10.1038/nrd3504













No comments:
Post a Comment